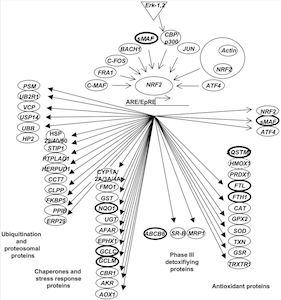
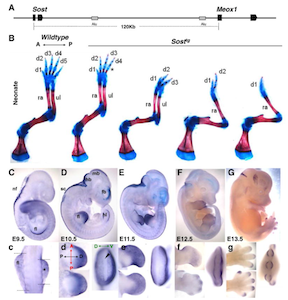
The Nrf2-Keap1 regulatory pathway regulates arsenic responsive genes
Known and novel arsenic responsive Nrf2 transcripts are shown. Empty circles depict unaffected target transcripts (not differentially expressed in lymphoblastoid cells). Bold and no continuous line circles indicate the level of increased gene expression as identified within this study. Bold line is 2–5 fold, no continuous line is more than 10 fold. Bold letters depict novel Nrf2-target genes identified by microarrays. PLoS One. 2014.